内吞体,作为细胞内囊泡转运的关键成员,可通过分裂、融合等形态改变精密地调控转运物质的分选,进而影响其胞内运输途径,包括进入溶酶体进行降解,抑或转运至细胞膜或反式高尔基体进行再循环利用。之前研究发现,内质网可通过与内吞体互作,从而调控内吞体的分裂,是否还有其它膜性细胞器参与调控该过程目前并不清楚。
2021年6月28日,清华大学生命学院贾顺姬副研究员(孟安明教授团队)联合中科院生物物理所李栋研究员实验室在Nature Cell Biology上发表了题为“高尔基体分泌小泡通过促进磷脂酰肌醇转换介导内吞体分裂(A Golgi-derived vesicle potentiates PtdIns4P to PtdIns3P conversion for endosome fission)”的研究论文。该论文报道了高尔基体,通过出芽小泡的方式调控内质网所介导的内吞体分裂,证实了细胞内不同膜器系统之间的高度协同互作。
在本研究中,研究者通过高分辨率成像结合电镜观察,鉴定了一种来源于高尔基体并高表达SEC14L2的小泡,名为SEC14L2囊泡。利用化学抑制剂BFA处理细胞、阻断高尔基体小泡形成时,内质网所介导的内吞体分裂过程严重受阻,内吞体呈现变大、拉管等形态异常,暗示高尔基体在内吞体分裂过程中发挥关键调控功能。为实时观察SEC14L2囊泡、内质网和内吞体三者之间的动态互作及形态变化,研究者利用高分辨快速成像系统GI-SIM发现,SEC14L2囊泡与内质网和内吞体均存在频繁的动态互作,并且被特异地招募至内质网所介导的内吞体分裂位点处。与此同时,在内吞体分裂前,SEC14L2囊泡促进内吞体中PtdIns3P的大量积累和PtdIns4P的去除。只有当内吞体完成了PtdIns4P向PtdIns3P的转换之后,内吞体才能实现正常的分裂过程。在机制上,研究者发现,该高尔基体来源囊泡上的SEC14L2蛋白发挥了关键作用。通过体外重构的方法,研究者证实SEC14L2蛋白,一方面可以直接结合PtdIns3P和PtdIns4P,介导它们在不同的脂质体之间转运;另一方面, 可以与PI3K结合,促进其激酶活力,用于PtdIns3P的合成。当敲除细胞内SEC14L2蛋白后,内吞体中的PtdIns3P含量显著下调,而PtdIns4P含量则急剧上升,同时会引起内吞体分裂过程受阻,内吞体体积增大。通过回补实验,研究者发现,在SEC14L2敲除的细胞中,内吞体的分裂异常可以很好的被野生型SEC14L2所挽救,而不能被PtdIns3P结合缺陷型的SEC14L2(SEC14L2-M5)所拯救,证明SEC14L2的磷脂酰肌醇结合能力对于SEC14L2囊泡调控内吞体的分裂是必须的。值得一提的是,在本文投稿过程中,Science杂志也发表了一篇高尔基体来源小泡调控线粒体分裂的文章,充分证明高尔基体来源小泡对于细胞内膜器系统稳态的重要性,而关于两种小泡是否为相同属性则需进一步验证。
清华大学生命科学学院贾顺姬副研究员和中科院生物物理所李栋研究员为该论文的通讯作者。清华大学生命学院博士后龚波和生物物理所博士后郭玉婷为该论文的共同第一作者。清华大学生命学院博士生丁诗卉、蛋白质中心脂质代谢平台刘晓蕙副研究员、清华大学生命学院孟安明教授为本文的共同作者。
该研究受到国家自然科学基金委重大研究计划(基金号:91754112、91754202),中国科技部重点研发计划(基金号:2019YFA0801403),国家自然科学基金委青年科学基金(基金号:31801130、32000480)和博新计划(基金号:BX20190355)等基金资助。
原文链接:https://www.nature.com/articles/s41556-021-00704-y
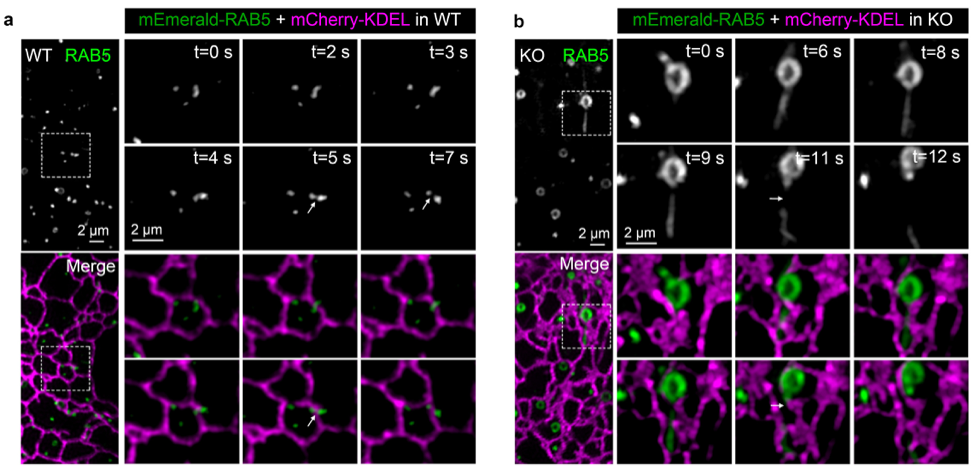
图1. 与野生型的COS7细胞(a)相比,SEC14L2敲除的细胞(b)表现为内吞体分裂延迟及体积增大。

图2. 高尔基体分泌的SEC14L2小泡与内质网协同调控内吞体分裂的模式图。SEC14L2小泡通过与内质网和内吞体之间形成膜相互作用,促进内吞体上PtdIns4P向PtdIns3P转换,从而促进内吞体分裂。

