阿尔兹海默症(Alzheimer’s disease)简称AD,亦称老年痴呆症,是当今世界范围内患病最广泛,病情最严重的神经退行性疾病。临床表现为记忆缺失、运动功能障碍,最后脑功能严重丧失直至死亡。随着年龄的增加,该疾病的发病概率逐渐增大,统计表明,在65岁以上人群中,其发病率高达10%,在85岁以上人群中,发病率更是达到30-50%。然而到目前为止,该疾病没有任何特效药物,预防方法和治疗手段都严重不足,给病人及其家人造成极大的痛苦,也为社会带来沉重的负担。
AD病人的特异性标志物之一是脑组织中出现的淀粉样斑块沉积,它是由淀粉样蛋白前体蛋白(APP, amyloid precursor protein)被γ-分泌酶(γ-secretase)切割后产生的Aβ多肽逐渐聚集形成的。γ-分泌酶是由四种亚基Presenilin1(PS1)、Pen-2、Aph-1和Nicastrin(NCT)共同组成的膜内蛋白酶复合物。遗传学证据表明,可遗传的家族性AD所携带的基因突变中有200多个突变位于编码γ-分泌酶活性亚基PS1的基因上。一直以来,抑制γ-分泌酶的切割活性,减少淀粉样斑块沉积都是一个重要的治疗AD的策略,很多针对AD药物的研发直接以γ-分泌酶作为靶点,希望通过调节其活性来治疗疾病,一些药物也曾进入临床II期或III期试验。非常遗憾的是,这些药物竟无一例外地宣告失败,AD药物研发领域倍受挫折。
礼来公司研发的小分子药物Semagacestat是最为著名的案例:Semagacestat是第一个进入临床三期试验的γ-分泌酶抑制剂,该小分子药物由于其前期试验的显著效果倍受关注;但是临床三期试验结果显示,该药物并未延缓疾病发展,而且还会导致患者认知能力和日常活动能力减弱。除此之外,Semagacestat还会提高病人患皮肤癌的风险,这很有可能是由于这种抑制剂不仅抑制APP的切割,还抑制了γ-分泌酶另外一种重要底物—Notch的切割,因此合理地设计对APP和Notch具有底物选择性的γ-分泌酶抑制剂具有重要的价值。然而一直以来γ-分泌酶与其小分子药物的原子分辨率结构的缺失使得进一步的药物设计与优化面临着很大的困难。
2020年12月28日,施一公教授团队在线发表“小分子药物抑制及调节γ-分泌酶的结构基础”(Structural Basis of γ-Secretase Inhibition and Modulation by Small Molecule Drugs)的文章[1],第一次报道了γ-分泌酶结合三种小分子抑制剂(Gamma-Secretase Inhibitor, GSI)和一种调节剂(Gamma-Secretase Modulator, GSM)的四个原子分辨率冷冻电镜结构,阐明了γ-分泌酶识别不同种类抑制剂及调节剂的分子机理。科研人员通过与2015年解析的γ-分泌酶的结构,以及2019年γ-分泌酶分别结合底物Notch与APP两种状态进行比较分析,首次完整展现了γ-分泌酶结合底物与药物的全过程,为了解γ-分泌酶活性调节机制提供了前所未有的精准蓝图,也将极大地推进下一代γ-分泌酶抑制剂及调节剂的设计与优化。

失败是成功之母,只有明白了为什么失败,才能积累经验,通向成功。在该研究中,施一公教授课题组首先解析了γ-分泌酶与Semagacestat的3.0 Å分辨率结构。Semagacestat在γ-分泌酶中与底物结合γ-分泌酶所形成的β-strand占据着相同的位置,对底物进入γ-分泌酶活性中心形成了位阻。值得一提的是,底物Notch与APP所形成的β-strand均会被Semagacestat阻碍,这一结果成功地解释了Semagacestat抑制γ-分泌酶活性的机制,也一目了然地说明了Semagacestat产生副作用的机制。
接下来的研究中,解释对APP和Notch具有一定选择性的抑制剂如何结合γ-分泌酶,对于开发或优化特异性抑制剂意义重大。施一公教授课题组选择了进入到临床II期由BMS公司研发的Avagacestat进行研究,成功解析了γ-分泌酶结合Avagacestat的3.1 Å的结构。出乎意料的是,Avagacestat也占据着底物的β-strand的位置,但是在与PS1的具体相互作用方面与Semagacestat有所不同。Avagacestat主要与PS1的疏水残基存在相互作用,只形成了一个氢键,而Semagacestat可以形成四个氢键。结合Avagacestat与结合Semagacestat后的PS1的构象有一定区别,呈现出更为“松散”的状态,Avagacestat的底物选择性或许和这些相互作用形式的不同有关。
Semagacestat和Avagacestat都是γ-分泌酶抑制剂中的非过渡状态类似物(non-transition state analog,non-TSA),除此之外,还有过渡状态类似物(transition state analog,TSA)抑制剂。为了更全面的阐述所有类型抑制剂的作用机理,他们进一步解析了γ-分泌酶结合TSA抑制剂L685,458的的结构。在γ-分泌酶研究领域,L685,458具有独特的地位,因为很多用于研究γ-分泌酶构象变化的化学工具都是L685,458的衍生物。意料之中,L685,458的一部分也具有与前面两种抑制剂相同的空间占位。不同之处在于,L685,458 还可以与γ-分泌酶的两个天冬氨酸催化残基存在直接相互作用,清楚的捕捉到了天冬氨酸水解酶催化过程中形成“正四面体”中间态的构象。
至此,该研究成功揭示了三种不同的γ-分泌酶抑制剂的作用机理,分析了其共同点,更重要的是揭示了三者之间的区别,为未来优化和设计具有底物特异性的抑制剂奠定了坚实基础。
然而抑制机理的研究只是γ-分泌酶活性调节的一个环节,在以γ-分泌酶为靶点的小分子药物设计领域,还有另一类不容忽视的小分子,即γ-分泌酶活性调节剂(GSMs)。γ-分泌酶调节剂(GSMs)在AD药物的研发方面也具有很大的潜力。与GSI不同的是,GSMs并不完全抑制γ-分泌酶切割活性,而是一定程度上促进毒性较强的Aβ42的进一步切割,形成更为安全的Aβ38等更短的多肽,缓解淀粉样斑块的形成。因此GSMs或许可以辅助GSIs以达到更优的切割效率和底物选择性。E2012是一种杂环类化合物,它可以和L685,458同时结合γ-分泌酶达到更高的抑制效果。该研究中展示了γ-分泌酶和E2012、L685,458的冷冻电镜结构,分辨率高达2.6 Å。E2012像一把匕首,一侧插入到PS1和NCT共同形成的结合空腔中,这也是首次明确了γ-分泌酶结合GSM的位点。E2012结合后可以和PS1的loop1形成氢键,loop1与γ-分泌酶的底物也存在相互作用。对比底物结合的构象,E2012空间上对底物跨膜螺旋的一侧具有一定的位阻效应,但不完全重合,这与抑制剂是有明显区别的。这种位阻效应,客观上构成了底物跨膜螺旋两个方向的运动趋势,为解释E2012促进Aβ42切割成Aβ38的别构调节机制提供了结构基础。
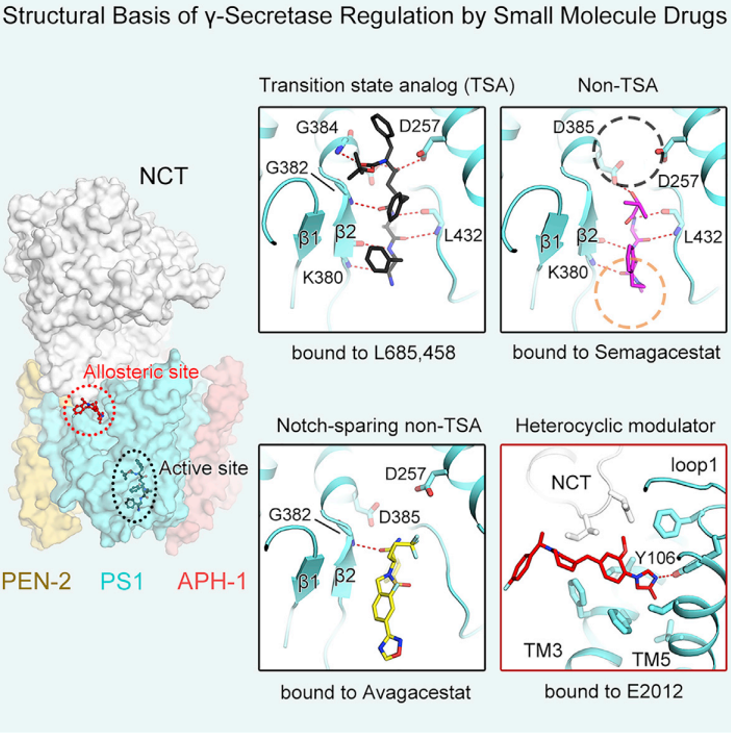
γ-分泌酶结合三种小分子抑制剂(Semagacestat、Avagacestat和L685,458)和一种调节剂(E2012)的原子分辨率冷冻电镜结构。三种小分子抑制剂结合在相同的位置,占据底物β-strand的位置,与PS1的β-strand相互作用。调节剂E2012结合在另外一个別构调节位置。
施一公教授实验室一直以来都将揭示阿尔兹海默症的发病机理作为重点研究方向,其中与AD相关的重要蛋白γ-分泌酶的三维结构及其作用机理的解析是一项重要的环节。2012年,施一公教授及其团队在Nature上发表文章报道了PS1的同源蛋白PSH的晶体结构[2]。2014-2015年,先后在Nature报道了分辨率为4.5 Å以及3.4 Å人源γ-分泌酶的三维结构,第一次报道了人源γ-分泌酶的冷冻电镜三维重构结构,并分析了γ-分泌酶中催化亚基PS1上的致病突变[4]。2019年,经过近五年的不懈努力,分别在Nature和Science发表论文报道了γ-分泌酶结合重要底物Notch和淀粉样前体蛋白APP的冷冻电镜结构,分辨率达到了2.7 Å和2.6 Å,这两篇文章为理解γ-分泌酶特异性识别并切割底物的分子机制提供了重要基础[5,6]。此次研究是在之前研究基础上γ-分泌酶结构领域的又一次进步与突破,为开发以γ-分泌酶为靶点的特异性药物提供了极为重要的基础。
施一公教授是该研究论文的通讯作者,该文章发表在国际著名学术杂志《细胞》(Cell)上。原清华大学结构生物学高精尖创新中心卓越学者,现中国农业大学教授杨光辉、清华大学生命学院助理研究员周瑞、清华大学生命学院三年级博士生郭雪飞为论文的共同第一作者。清华大学高精尖创新中心的研究员闫创业博士参与了本项工作,清华大学冷冻电镜平台的雷建林博士对数据收集进行了指导。国家蛋白质科学中心(北京)清华大学冷冻电镜平台和清华大学高性能计算平台分别为该研究的数据收集和数据处理提供了支持,冷冻电镜平台的李晓敏博士、杨帆博士对数据收集提供了帮助。北京市结构生物学高精尖创新中心(清华)、科技部国家重点研发计划、国家自然科学基金、浙江省重点研发计划为该研究提供了经费支持。
原文链接
1. Yang et al., Structural Basis of γ-Secretase Inhibition and Modulation by Small Molecule Drugs, Cell (2021); https://www.cell.com/cell/fulltext/S0092-8674(20)31621-4
相关论文
2. Li, X., Dang, S., Yan C., Gong, X., Wang, J., Shi, Y. (2013). Structure of a presenilin family intramembrane aspartate protease. Nature 493, 56–61
3. Lu, P., Bai, X.C., Ma, D., Xie, T., Yan, C., Sun, L., Yang, G., Zhao, Y., Zhou, R., Scheres, S.H.W., and Shi, Y. (2014). Three-dimensional structure of human gamma-secretase. Nature 512, 166-170.
4. Bai, X.C., Yan, C., Yang, G., Lu, P., Ma, D., Sun, L., Zhou, R., Scheres, S.H.W., and Shi, Y. (2015b). An atomic structure of human gamma-secretase. Nature 525, 212-21
5. Yang, G., Zhou, R., Zhou, Q., Guo, X., Yan, C., Ke, M., Lei, J., and Shi, Y. (2019). Structural basis of Notch recognition by human gamma-secretase. Nature 565, 192-197.
6. Zhou, R., Yang, G., Guo, X., Zhou, Q., Lei, J., and Shi, Y. (2019). Recognition of the amyloid precursor protein by human gamma-secretase. Science 363.

